
Índice de contenidos
El Sistema del Complemento
El término complemento, definido por el famoso inmunólogo Paul Ehrlich como la actividad del suero que completa la acción de los anticuerpos, hace referencia a un conjunto de proteínas del suero que colaboran, tanto con el sistema inmune innato, como con el adaptativo para eliminar los patógenos en sangre y tejidos. De la misma forma que los componentes sanguíneos intervienen en la coagulación, las proteínas del complemento interaccionan con otras cascadas catalíticas.
Actualmente, sabemos que la acción del complemento es el resultado de interacciones entre un complejo grupo de más de 30 glicoproteínas. La mayoría de los componentes del complemento son sintetizados por hepatocitos, aunque algunos se producen por otros tipos celulares, como los monocitos, macrófagos, fibroblastos y células epiteliales del tracto gastrointestinal y genitourinario.
Los componentes del complemento se pueden clasificar en siete categorías funcionales (Figura 1):
(1) Las rutas del complemento se inician por proteínas como el C1q que se unen a patógenos o bien directamente o por anticuerpos o por proteínas específicas de patógeno.
(2) Mediadores enzimáticos que activan otras enzimas que generan proteínas centrales de la cascada del complemento, las C3 y C5 convertasas, las cuales rompen C3 y C5, liberando componentes activos que median todas las funciones del complemento, incluyendo (3) opsonización, (4) inflamación y (5) la generación del complejo de ataque a la membrana (MAC).
Las proteínas efectoras del complemento pueden marcar un complejo antígeno-anticuerpo por fagocitosis (opsoninas), iniciar inflamación (anafilotoxinas) o unirse a un patógeno y formar el MAC. A menudo, estos efectores actúan a través de (6) receptores del complemento en células fagocíticas, granulocitos o eritrocitos.
(7) Asimismo, proteínas reguladoras limitan los efectos del complemento mediante la promoción de su degradación o prevención de su unión a las células del huésped.
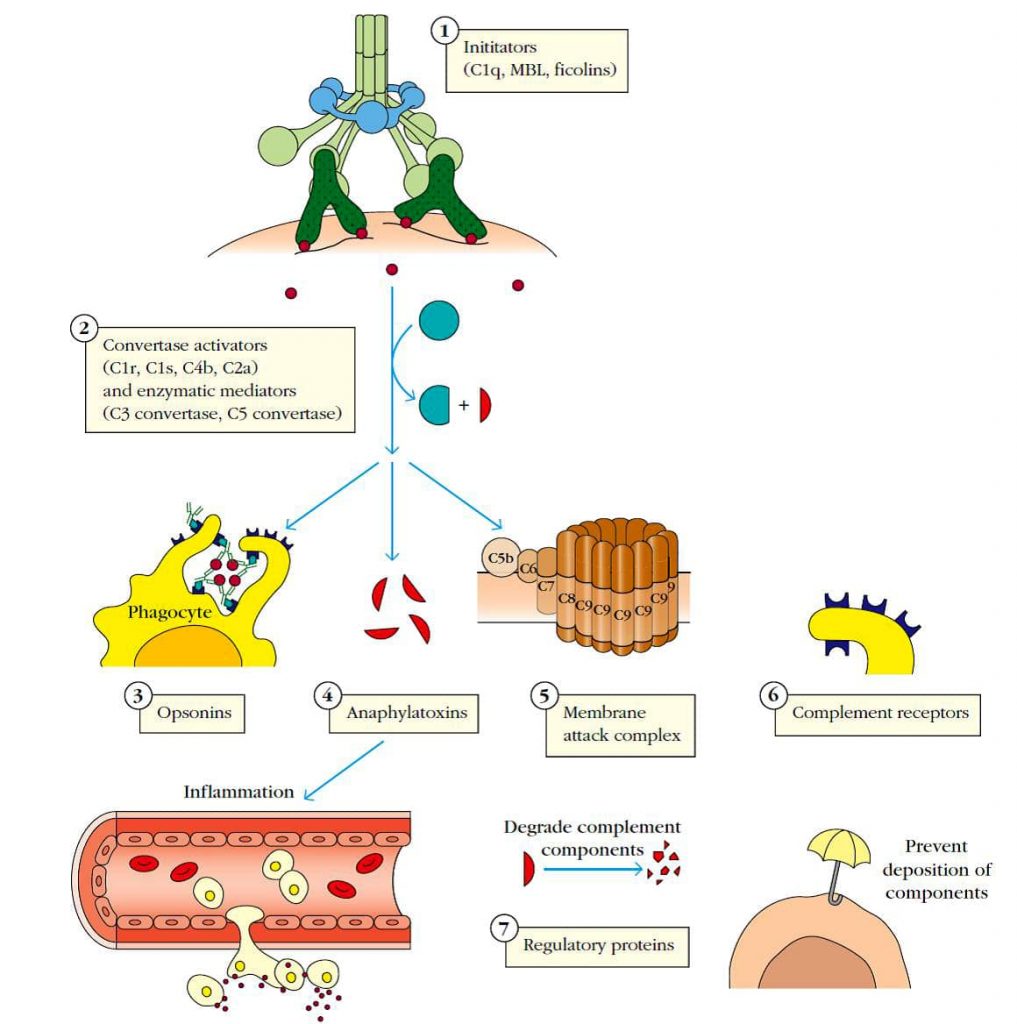
Figura 1: Proteínas que participan en el sistema del complemento1.
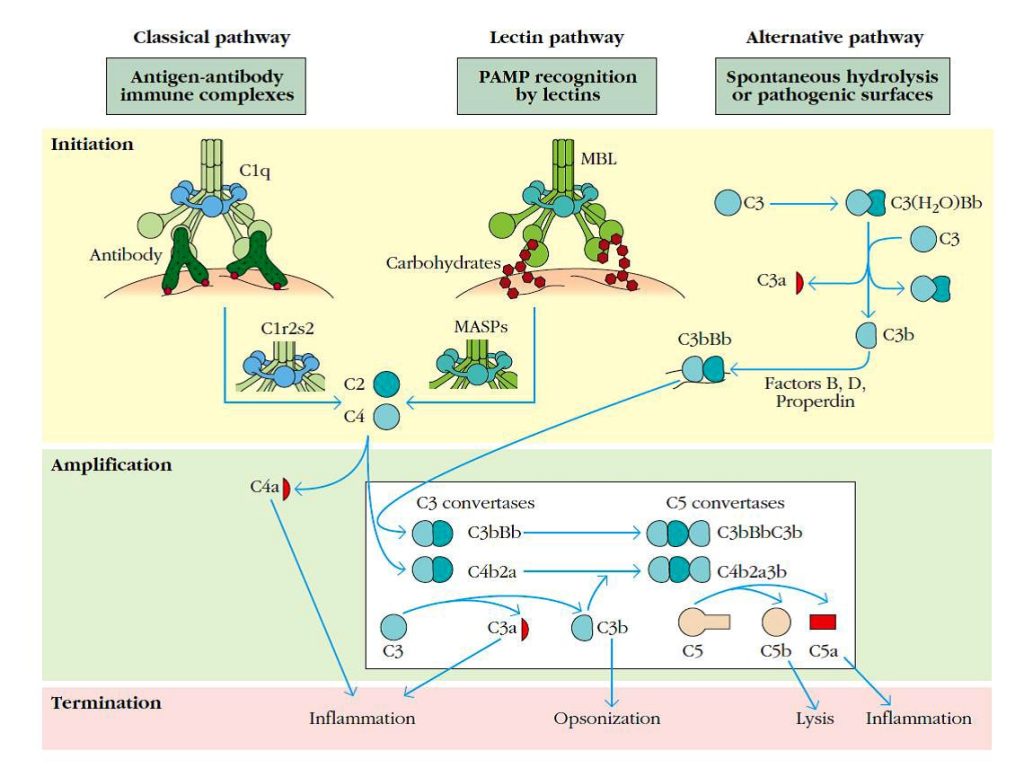
El sistema del complemento se puede activar por tres vías. La ruta clásica se inicia cuando C1q se une a complejos antígeno-anticuerpo. El componente enzimático C1r de C1 se activa y rompe C1s, el cual a su vez rompe C4 en C4a (una anafilotoxina) y C4b. C4b se une a la membrana y a C2, el cual se rompe por C1s para formar C2a y C2b. Así, C2a permanece unido a C4b, formando la convertasa C3 de la ruta clásica (C4b2a). En la ruta de las lectinas, las lectinas de unión a manosa (MBL) se unen específicamente a carbohidratos conservados en patógenos, activando serín proteasas asociadas a MBL (MASPs).
Las MASPs rompen C2 y C4 generando la convertasa C3 como en la vía clásica.
En la ruta alternativa para la activación del sistema del complemento, C3 se somete a una hidrólisis espontánea para dar C3(H2O), el cual se une al factor B del suero. En esta unión, B se rompe por el factor D y el complejo C3(H2O)Bb resultante forma la convertasa C3 en fase fluida. Algunas C3b, liberadas después de la ruptura de C3 por este complejo, se unen a la superficie de los microorganismos.
Así, se une al factor B, el cual se rompe por el factor D, formando la convertasa C3 de la ruta alternativa, C3bBb. Este complejo se estabiliza por properdina.
Finalmente, las convertasas C5 se forman por la adición de un fragmento C3b para cada convertasa C3. Las C5 convertasas se rompen en C5a y C5b, mediadores inflamatorios que inician la formación del complejo de ataque a membrana (Figura 2), con la adición de los componentes C6, C7, C8 y C9 al componente C5b. Este complejo induce la formación de grandes poros en la membrana, alterando su integridad osmótica y provocando la muerte celular.
La molécula C1q
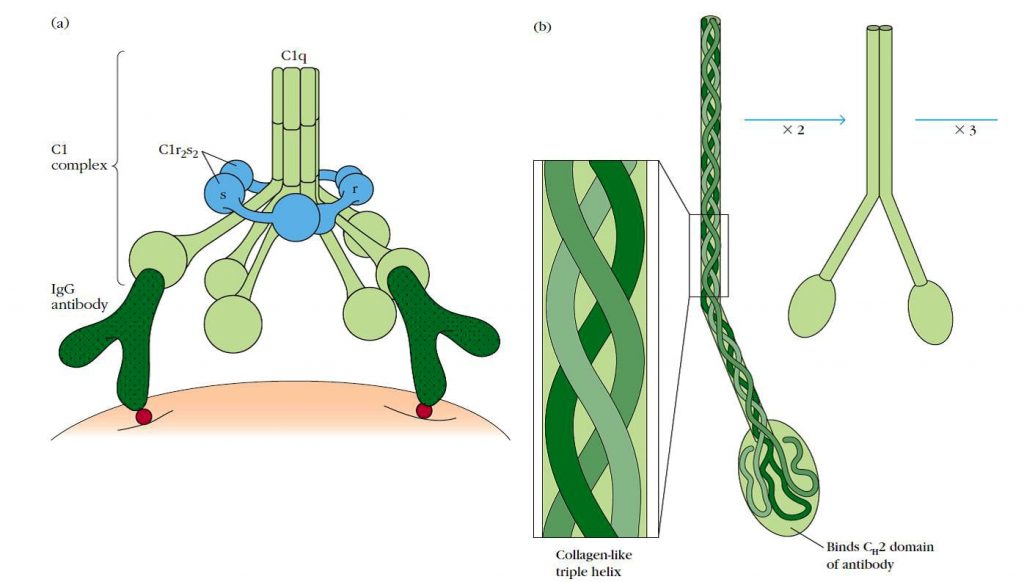
La formación de un complejo antígeno-anticuerpo induce cambios conformacionales en la porción Fc del anticuerpo. Este cambio conformacional expone un sitio de unión para el componente C1 del complemento. En suero, C1 existe como un complejo macromolecular de cinco proteínas que consiste en una molécula C1q y dos moléculas de cada una de las serín proteasas, C1r y C1s, (C1qr2s2), estabilizado por Ca2+.
La molécula C1q está compuesta por 18 cadenas polipeptídicas que se asocian para formar 6 brazos de triple hélice de colágeno, los extremos de los cuales se unen al dominio CH2 de la unión del antígeno al anticuerpo (Figura 3).
Cada complejo macromolecular debe unirse por sus cabezas globulares C1q a al menos dos sitios Fc para que se produzca una interacción estable C1-anticuerpo.
Estas cabezas globulares son similares a dominios encontrados en la superfamilia de proteínas del factor de necrosis tumoral (TNF). Además, se han asociado a C1q otras funciones relacionadas con el desarrollo de problemas del sistema nervioso central que suceden con la edad y quizá la regulación del crecimiento tumoral2.
Funciones del Sistema del Complemento
La actividad del sistema del complemento puede dividirse en tres áreas:
- Defensa innata frente a las infecciones: lisis de bacterias y membranas celulares a través del MAC, opsonización por unión covalente de C3b y C4b e inducción de inflamación y quimiotaxis por anafilotoxinas como C3a, C4a y C5a.
- Interconexión entre la inmunidad innata y adaptativa: aumentando la respuesta de los anticuerpos, favoreciendo la memoria inmunológica y la presentación antigénica. Las proteínas del complemento se unen a receptores en la superficie de las células presentadoras de antígenos, células T y B, induciendo la producción de citoquinas e interviniendo en la activación de estas células.
- Intervención en la fase de contracción de la respuesta inmune: eliminando complejos inmunes de tejidos y células apoptóticas gracias al componente C1q e inducción de células T reguladoras. La unión del componente C1q del complemento a células apoptóticas, cuerpos apoptóticos e inmunocomplejos provoca su opsonización con C3b y la subsecuente fagocitosis.
El sistema de proteínas reguladoras del complemento garantiza que la activación del complemento no se produzca en la superficie de las células huésped mediante la desactivación de los componentes del complemento en la superficie de dichas células y asegurando que las proteínas reguladoras se unan específicamente a esas células y no a la membrana de microorganismos celulares.
Virus, bacterias, hongos y parásitos han desarrollado una gran variedad de estrategias para evadir al sistema del complemento, incluyendo la mimetización con proteínas reguladoras, interferencia con las interacciones entre anticuerpo y componentes del complemento o entre proteínas de las rutas del complemento o por destrucción de sus componentes3.
Relación de anticuerpos anti-HLA y C1q
Actualmente, la presencia de anticuerpos anti-HLA está asociada con un aumento de rechazo y disminución de la supervivencia de los trasplantes. Los anticuerpos se miden en ensayos semicuantitativos, donde se estratifican los resultados en función de la intensidad media de fluorescencia (MFI). Además de esto, se valoran otras propiedades como la capacidad de unión del complemento o el perfil de las subclases de IgG1-4.
Muchos artículos han mostrado que el C1q se correlaciona significativamente con un alto riesgo de rechazo mediado por anticuerpos o pérdida del injerto.
Los anticuerpos específicos de donante (DSA) C1q+ son clínicamente más relevantes que DSA IgG+. Aproximadamente el 50% de los anticuerpos IgG+ son también C1q+4, pudiendo categorizarse los anticuerpos en base a su habilidad para fijar el complemento. El ensayo C1q puede ser utilizado pre-trasplante junto con el screening de anticuerpos de rutina y post-trasplante para monitorizar la eficacia de los tratamientos de rechazo.
Parece que hay una relación directa entre la capacidad de unirse al sistema del complemento y la fuerza de esos anticuerpos anti-HLA5, 6, 7. Aunque quedan todavía aspectos por resolver, ya que anticuerpos con valores bajos de MFI (<5000) pueden unirse fuertemente a C1q y viceversa.
De entre todos los isotipos de inmunoglobulinas, IgG es considerado el principal efector del rechazo humoral a través de la activación de la vía del complemento. Sin embargo, las 4 subclases presentan diferentes propiedades8, 9, 10, fijando el complemento en el siguiente orden: G3>G1>>G2>G4. La presencia de una mezcla de subclases es más común en anticuerpos con alta especificidad de unión a C1q10.y una respuesta inmune más avanzada, estimulada por una exposición a antígeno más larga y prolongada.
Pero, no está clara la asociación de la capacidad de unión al complemento con la diferente composición de subclases11, ya que la proporción de IgG1/G3 es similar en anticuerpos fijadores y no fijadores de complemento.
Sin embargo, las subclases de IgG no pueden usarse en determinados casos como en el caso de pacientes desensibilizados con inmunoglobulinas intravenosas u otros fármacos basados en anticuerpos humanos, puesto que también contienen subclases de IgG. Por tanto, es muy interesante y necesario llevar a cabo el ensayo C1q para estar seguros del estado de fijación del complemento por los anticuerpos.
Por tanto, es muy interesante y necesario llevar a cabo el ensayo C1q para estar seguros del estado de fijación del complemento por los anticuerpos.
Reto para el futuro
Saber en qué momento realizar un trasplante, con qué donante, cuándo y por cuánto tiempo desensibilizar y cuándo aumentar o eliminar la inmunosupresión son los objetivos a lograr.
Hoy en día, parece que una combinación de ensayos C1q e IgG (MFI/subclases/titulación) podría resultar el escenario más informativo a la hora de evaluar el riesgo de un paciente antes y después del trasplante.
Bibliografía
1.- Owen JA, Punt J, Strandford SA, Jones PP. Kuby Immunology. 7th Edition (2013).
2.- Reid KBM. Complement Component C1q: Historical Perspective of a Functionally Versatile, and Structurally Unusual, Serum Protein. Front Immunol 10;9:764 (2018).
3.- Blom, A. M., T. Hallstrom, and K. Riesbeck, K. Complement evasion strategies of pathogens—acquisition of inhibitors and beyond. Molecular Immunology 46:2808–2817 (2009).
4.- Chen G, Sequeira F, Tyan DB. Novel C1q assay reveals a clinically relevant subset of human leukocyte antigen antibodies independent of immunoglobulin G strength on single antigen beads. Hum Immunol 72:849–858 (2011).
5.- Tambur AR, Herrera ND, Haarberg KM, Cusick MF, Gordon RA, Leventhal JR, et al. Assessing antibody strength: comparison of MFI, C1q, and titer information. Am J Transplant. 15:2421–30 (2015).
6.- Molina J, Navas A, Aguera ML, Rodelo-Haad C, Alonso C, Rodriguez-Benot A, et al. Impact of preformed donor-specific anti-human leukocyte antigen antibody C1q-binding ability on kidney allograft outcome. Front Immunol. 8:1310 (2017).
7.- Zeevi A, Lunz J, Feingold B, et al. Persistent strong anti-HLA antibody at high titer is complement binding and associated with increased risk of antibodymediated rejection in heart transplant recipients. J Heart Lung Transpl 32:98–105 (2013).
8.- Hickey MJ, Valenzuela NM, Reed EF. Alloantibody generation and effector function following sensitization to human leukocyte antigen. Front Immunol. 7:30 (2016).
9.- Valenzuela NM, Hickey MJ, Reed EF. Antibody subclass repertoire and graft outcome following solid organ transplantation. Front Immunol. 7:433 (2016).
10.- Valenzuela NM, Schaub S. The biology of IgG subclasses and their clinical relevance to transplantation. Transplantation. 102:S7–13 (2018).
11.- Navas A, Molina J, Agüera ML, Guler I, Jurado A, Rodríguez-Benot A, Alonso C, Solana R. Characterization of the C1q-Binding Ability and the IgG1-4 Subclass Profile of Preformed Anti-HLA Antibodies by Solid-Phase Assays. Front Immunol. 10:1712 (2019).
Artículos relacionados
- Inmunoterapia asociada a HLA
- AT1R y ETAR, receptores de angiotensina y endotelina
- El deporte y sus beneficios en el sistema inmunológico
- Moléculas de histocompatibilidad no clásicas: MICA
- Influencia de los sistemas HLA y HNA en la susceptibilidad, protección inmunológica y éxito de tratamientos novedosos frente a COVID-19
- Anticuerpos HLA en el trasplante hepático
- Crossmatch Virtual
- La respuesta inmune frente al cáncer